Archives
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-11
- 2018-10
- 2018-07
-
In contrast T bet downregulated the
2019-08-02
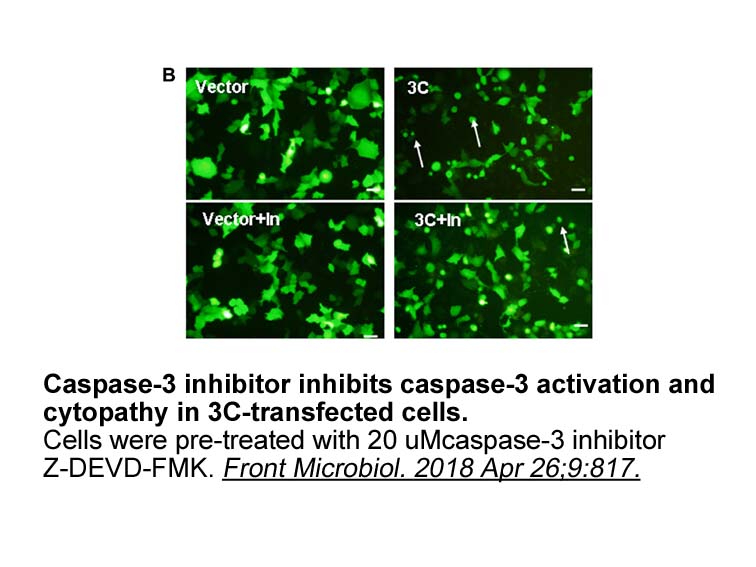
In contrast, T-bet downregulated the expression of Tfh functional molecules, CD40L, ICOS, and IL-21; accordingly, T-bet overexpression abrogated the helper function of IL-12-stimulated CD4+ T cells. Thus, T-bet seems more involved in the downregulation of the helper functions than in the differentia
-
Through an unknown mechanism RING
2019-08-02
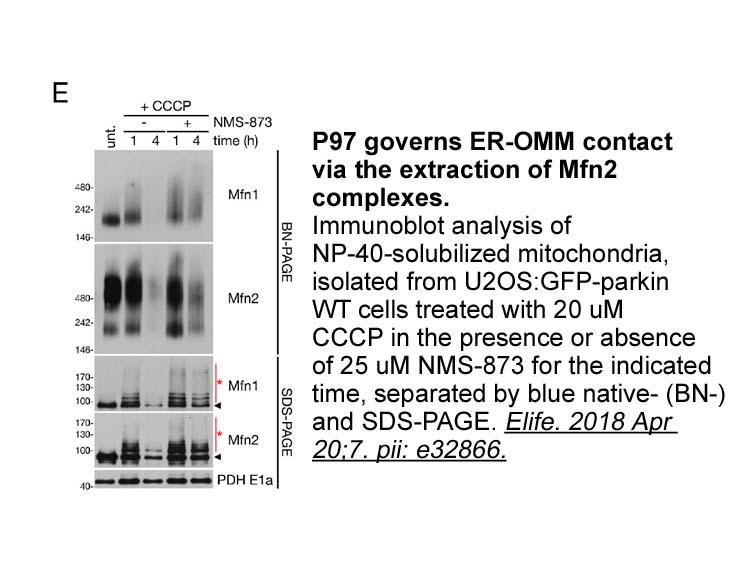
Through an unknown mechanism, RING/Ubox-type E3 dimers and the N-terminal tail have been shown to function as a modulator of full E3 ubiquitin ligase activity [27]. LRSAM1 promotes a significant enhancement in the formation of the high-molecular-weight products or E3 activity when the RING domain is
-
cyp3a inhibitors Compounds that interact with MDR
2019-08-02
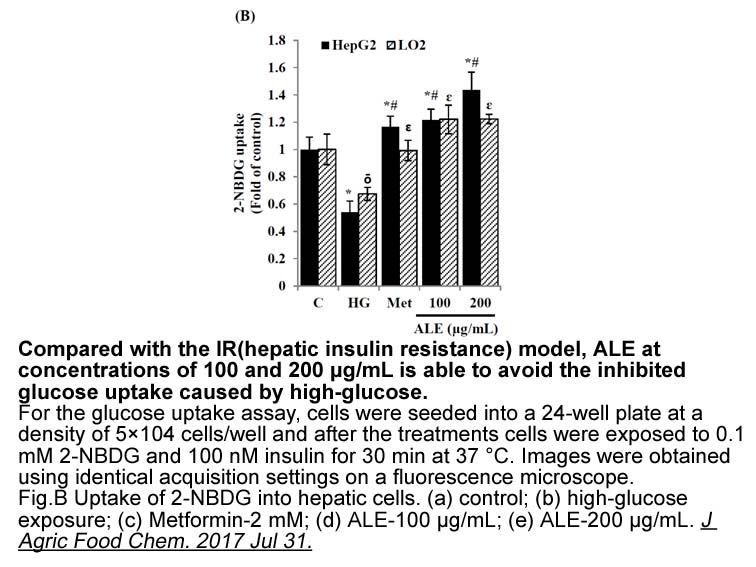
Compounds that interact with MDR1 can do so by different mechanisms. Verapamil is known to modulate drug resistance by acting as a competitive MDR1 substrate [36]. Interestingly, NU7441 has similar growth inhibitory activity in the sensitive and resistant cyp3a inhibitors and there was no observed
-
br Enzymes involved in microbial steroid degradation
2019-08-02
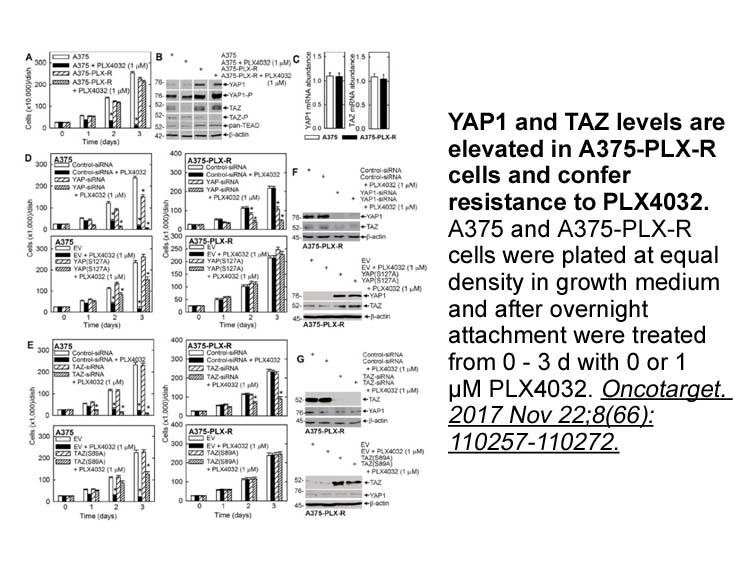
Enzymes involved in microbial steroid degradation are generally not expressed constitutively, but they are upregulated depending on which steroid substrates are present [17,81]. Thus, a cell-free extract prepared from testosterone-adapted C. testosteroni ATCC 11996 cells displayed a 1(2)-dehydroge
-
For immobilization of enzymes natural polymers
2019-08-01
For immobilization of enzymes, natural polymers depict several outstanding features as support. These materials are inert, non-toxic, biodegradable and biocompatible. As well as, they can be treated with different functional groups easily by chemical reactions that occur under gentle conditions in a
-
Next we evaluated neutralization of Jc Jc N A Jc
2019-08-01
Next, we evaluated neutralization of Jc1, Jc1-N534A, Jc1-ΔHVR1 and Jc1-ΔHVR1-N534A by sera from individuals chronically infected with HCV GT1 and GT2 (Fig. 4A-C). Like the results of neutralization with monoclonal Miconazole where and CD81-LEL (Fig. 3), the sera poorly neutralized Jc1. Neutralizati
-
br Structural and biochemical features of pol
2019-08-01
Structural and biochemical features of pol X family polymerases The X-family DNA polymerase (polX) is specialized in DNA repair. This family is composed of four different DNA polymerases: pol β, pol λ, pol μ and TdT. Only three members of the polX family possess an N-terminal BRCT (BRCA1 carboxy-
-
br Funding br Conflict of interests br Introduction DAG is
2019-08-01
Funding Conflict of interests Introduction DAG is generated by the hydrolysis of phosphatidylinositol-4,5-bisphosphate (PtdIns(4,5)P2) by PtdIns-specific phospholipase C (PLC) enzymes [1]. Remaining in the membrane, it binds proteins with cysteine-rich, C1 domains, and activates several of
-
br Conflicts of interest br Introduction Diacylglycerols DAG
2019-08-01

Conflicts of interest Introduction Diacylglycerols (DAGs) and phosphatidic Compound C (PA) play fundamental roles in biology as basic components of membranes, intermediates in lipid metabolism, and secondary messengers in cellular signaling (Carrasco and Merida, 2007, Fang et al., 2001). Cell
-
In conclusion we have discovered two new
2019-08-01
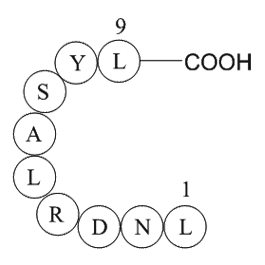
In conclusion, we have discovered two new derivatives ( and ) that are potent inhibitors of DHODH. H and C NMR spectroscopic data revealed that these compounds undergo ready isomerisation at room temperature in -DMSO, but the docking studies indicate that there is neither conformation nor configurat
-
Two key enzymes play important roles in
2019-08-01
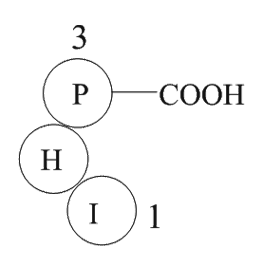
Two key enzymes play important roles in the bioconversion of xylitol from d-arabitol, namely d-arabitol dehydrogenase (ArDH) and NADH-dependent xylitol dehydrogenase (XDH), respectively (Qi et al., 2014). In an earlier study, we developed a high xylitol yield strain by expressing xdh in E. coli BL21
-
These cytokines except for TGF can also increase
2019-08-01
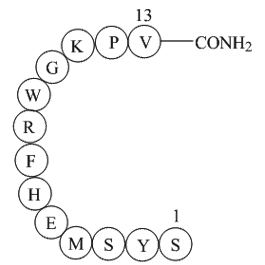
These cytokines, except for TGF-β can also increase CysLT1R mRNA expression. We further demonstrated that TGF-β and IL-13 were able to increase the proliferation of human BSMC in response to LTD4. BSMC proliferation was CysLT1R-dependent, given that Montelukast ablated this effect. Taken together, o
-
Orange spotted grouper Epinephelus coioides is
2019-08-01
Orange-spotted grouper (Epinephelus coioides) is a coral reef fish with a high commercial value in China and Southeast Asian countries. However, in recent years, various outbreaks of viral diseases have affected the grouper aquaculture industry (Wei et al., 2010). Specifically, Singapore grouper iri
-
More recently evidence suggests that dysregulation of CRF si
2019-08-01
More recently, evidence suggests that dysregulation of CRF signaling may also be implicated in panic disorder. For instance, polymorphisms in the CRF1, but not CRF2, gene have been associated with susceptibility to panic disorder in German and Japanese population samples (Ishitobi et al., 2012, Keck
-
Our present results support a major
2019-08-01
Our present results support a major role of EP1 receptors in peripheral heat sensitization and a smaller contribution to central heat sensitization but no contribution to mechanical sensitization. While the contribution of peripheral EP1 receptors to heat hyperalgesia is in good agreement with the a
16683 records 1030/1113 page Previous Next First page 上5页 10261027102810291030 下5页 Last page